Mechanosensitivity and Mechanotransduction: 4 (Mechanosensitivity in Cells and Tissues)
The modelling also decreased the capability for adhesion in these cells [ 90 ]. These changes are seen in alterations in division speed and development potential, and are probably connected with cytoskeleton reorganisation. Nevertheless, the primary levels of nonmuscle cellular responses to a change in external mechanical conditions are little studied and so the search for the mechanosensor will not end soon.
Results of numerous studies in conditions of real and simulated gravitational unloading testify that negative changes in various bodies and tissues are formed as a result of the action of microgravity. Skeletal muscles are especially vulnerable to a gravity-free state as the specialised organ that executes position and motor functions. A subject of many studies is the m. In addition, a decrease in the functionality of both the whole muscle [ , ] and single fibres [ ] also takes place.
Nevertheless, a substantive problem which is obstructing space development by humans, in particular flight to Mars, is the early readaptation period to gravity. Especially important is the reloading of muscle and cardiovascular system functionality, the acceleration of which is impossible without an understanding of the mechanism of development of adaptive changes. Differently directed changes in the external mechanical conditions of skeletal and cardiac muscles of rats can be achieved by means of a common method; antiorthostatic hindlimb suspension of animals, according to the Ilyin-Novikov method with Morey-Holton modifications [ ].
On the one hand, the antiorthostatic suspension of animals leads to hindlimb disuse, while on the other hand, it causes an increase in the mechanical load on cardiomyocytes. In addition, the orientation of skeletal muscle fibres and cardiomyocytes in the gravitational field will change. According to the scarce data connected with the response of skeletal muscles cells to gravity disuse in the literature, one of the first events to occur, after two days of antiorthostatic disuse of the hindlimbs of mice, is the accumulation of calcium ions in the soleus muscle [ , ].
Above, we described that the maximal calcium ion accumulation in the soleus muscle fibres of rats and a Mongolian gerbils happens after one day of gravity disuse. For the medial gastrocnemius and tibialis anterior muscles, this maximum was seen a short time later—after seven days of antiorthostatic suspension [ ]. The increase in the resting calcium level can lead to calpain activation [ , ] and the following destruction of the muscle fibre structure.
However, the means by which it is accumulated remain unknown. Probably, this effect happens via L-type calcium channels, but the mechanosensitive channels of the TRP family may also take part. In any case, the functioning of the channels incorporated in the membrane depends on the condition of the sarcolemma and the cytoskeleton connected with it. Moreover, we can suppose that the mechanical properties of cardiomyocytes will change, in other ways to that of skeletal muscle fibres, as the load on the cardiac muscle increases in microgravity conditions, while the load on the skeletal muscles of hindlimbs decreases.
It is rather difficult to perform a direct evaluation of the native state of the cortical cytoskeleton of muscle fibres. However, the definition of its mechanical properties transversal stiffness, to be exact can help in the analysis of its structural changes. In addition, the complicated sarcomere organisation of a muscle fibre enables us to suppose that the transversal stiffness of different parts of the sarcolemma Z-disk, M-line, and the part between them will differ from each other.
The differentiation of stiffness factors is of great interest, for example, in regarding the signalling role of different proteins of the costamere, which probably depends on its structure. The methodology based on this above concept enabled us to define the transversal stiffness both of a contractile apparatus and of a membrane with a cortical cytoskeleton [ 5 ]. It was shown that the transversal stiffness of different parts of the soleus muscle contractile apparatus in the relaxed, calcium activated, or rigor states decreases, over the course of gravity disuse.
It, in practical terms, does not change for the gastrocnemius muscle fibres and increases for tibialis anterior [ 8 , ]. Meanwhile, the transversal stiffness of the sarcolemma, with a cortical cytoskeleton in the relaxed state, decreases in all the muscles when under antiorthostatic disuse.
Furthermore, for rat cardiomyocytes, it seems, that under the conditions of antiorthostatic disuse, there was an increase in mechanical load early on as a result of hypovolemia. Consequently, the stiffness of the cortical cytoskeleton membrane increased: At reloading after antiorthostatic disuse, the stiffness of skeletal muscles fibres increased transversal stiffness of rat soleus muscle fibres: Changes in the transversal stiffness factors, both of skeletal muscles fibres and cardiomyocytes, under the conditions of antiorthostatic disuse and the following reloading, correlated with the content of nonmuscle isoforms of actin that form a cortical cytoskeleton in the membrane fraction.
They were also connected with differently directed changes in nonmuscle isoforms of the alpha-actinin content 1 and 4 of the membrane and cytoplasmic fractions [ 6 , ].
- .
- .
- The Oracle of the Seven Keys;
- Therons Tale: (a Witches of Etlantium novella)!
- Medo de Médico (Portuguese Edition).
For soleus muscle fibres, it was shown that the content of beta-actin in the cytoplasmic fraction did not change as a result of day suspension, nor in the following 3-day reloading. However the content of beta-actin in the membrane protein fraction decreased after disuse more than threefold when compared with the control level, but after three days of recovery, it did not differ from the control level, which correlated with the changes in sarcolemma transversal stiffness [ ]. Meanwhile, for cardiomyocytes the content of gamma-actin in the cytoplasmic protein fraction also remained at the level of the control during disuse and reloading.
However, the content of gamma-actin in the membrane protein fraction significantly increased during the first day of antiorthostatic suspension and continued to rise until the fourteenth day, showing similar dynamics to that of the transversal stiffness change.
Cell Mechanosensitivity: Mechanical Properties and Interaction with Gravitational Field
At the same time, the content of beta-actin in the cytoplasmic and membrane fractions did not change under the conditions of antiorthostatic disuse and the following reloading [ 6 ]. It should be noted that the increase of nonmuscle F-actin beta-actin content was noticed in cat cardiomyocytes during hypertrophy stimulation [ 8 ], although the content of gamma-actin was not defined during this experiment. The change in the content of nonmuscle actin isoforms, particularly in the membrane fraction, will lead to a structural change in the cortical cytoskeleton as well as changes in actin-binding protein content, particularly alpha-actinin-1 and alpha-actinin Considering data on the increase in the resting calcium levels of muscle fibres in gravity disuse, interest is drawn towards the calcium-sensitive actin-binding proteins-alpha-actinin-1 and alpha-actinin For rat cardiomyocytes under conditions of antiorthostatic disuse, the content of alpha-actinin-1 in the cytoplasmic fraction of proteins decreased after seven days of disuse, while in the membrane fraction it increased.
In the period of 3-day reloading after 14 days of antiorthostatic disuse, the content of alpha-actinin-1 in the membrane fraction decreased, while it increased in the cytoplasmic fraction. After seven days of recovery, both fractions were the same as the control level [ 6 ]. At the same time, in cardiomyocytes, the content of alpha-acitnin-4 in the membrane protein fraction increased during the first day, and starting from the third day its content increased from control levels in the cytoplasmic fraction as well.
During the reloading period the content of alpha-acinin-4 in the membrane fraction decreased to the control level. As for the cytoplasmic fraction, it also decreased, but did not reach the control level [ 6 ]. In addition, in the rat soleus muscle fibres under conditions of antiorthostatic disuse, the content of alpha-actinin-4 did not change in the cytoplasmic protein fractions.
In the membrane fraction, the content of alpha-acinin-4 decreased after 14 days of disuse, and increased after 3-day reloading, although it did not reach the same levels as the control [ ]. Very little is known about the role of nonmuscle isoforms of alpha-actinin in skeletal muscles cells or cardiomyocytes. It is known that alpha-acinin-1 is expressed in cardiomyocytes [ ] and in skeletal muscles cells, as well as alpha-actinin-4 in different stages of differentiation [ ]. Alpha-actinin-1 and alpha-actinin-4 are nonmuscle isoforms of alpha-actinin—a protein that belongs to the spectrin family [ ].
Mechanosensitivity and Mechanotransduction (Mechanosensitivity in Cells and Tissues)
They function via antiparallel homodimer binding of actin thread ends [ ]. In addition, alpha-actinin-4 connects the actin cytoskeleton to the membrane and facilitates interactions of the cortical cytoskeleton with cytoplasmic signalling proteins [ ]. Nevertheless, there is some data, showing that the increase in alpha-actinin-4 content in the cytoplasmic fraction is associated with a decrease of alpha-actinin-1, and with the formation of a cancerous pattern in fibroblasts [ ]. There are also results showing that the malignancy of cells, particularly lymphocytes, is followed by an increase of their stiffness, which was measured with the help of atomic force microscopy [ 11 ].
Therefore, the increase of cell stiffness is connected with the development of a cortical cytoskeleton, the decrease of alpha-actinin-1 and the increase of alpha-actinin-4 in the protein membrane fraction. Therefore, it can be supposed that the development of a cortical cytoskeleton will lead to the increase in alpha-actinin-1 and alpha-actinin-4 contents in membrane protein fractions.
Data obtained by Goffart et al. For muscle cells, it was shown that the functions of mitochondria can influence their shape, due to extension or compression of the membrane, that can be mediated by the cytoskeleton [ , ]. In addition, Milner et al. Moreover, these authors determined the intensity of cellular respiration and showed that both the level of oxygen uptake and the ADP dissociation constant markedly decreased in comparison with values in the control mice [ ].
Since, in the early stages of gravity disuse, the relative desmin content in m. These findings showed that cellular respiration decreased after three days of gravity disuse, achieved its minimum after seven days, and increased to the control level on the fourteenth day of simulated disuse [ ]. A three-day reloading period leads to some decrease in the cellular respiration, but recovery of the respiration parameters up to the control level was observed after seven days of readaptation [ ].
Meanwhile, according to Bigard et al. During work on cardiomyocytes, Saks et al. Our findings showed that the basal speed of cellular respiration of rat cardiomyocytes did not, in practical terms, change during antiorthostatic disuse; it did increase, but insignificantly, during the first day. Meanwhile on adding glutamate and malate to the medium, the respiration speed and maximum respiration speed significantly increased after just one day, and remained high for the whole period of disuse up to the fourteenth day.
- Cell Mechanosensitivity: Mechanical Properties and Interaction with Gravitational Field.
- .
- ;
- Morrhår under skalet (Swedish Edition);
Thus, after a 3-day-reloading after 14 days of antiorthostatic disuse, all indicated parameters significantly decreased in comparison to the control level [ 6 ]. However, Bigard et al. The discrepancy between our results and those of Bigard et al. The increase in the relative content of desmin in rat cardiomyocytes under conditions of antiorthostatic disuse [ 6 ], which is necessary for the definition of mitochondria localisation and regulation of the permeability of their membranes, in combination with data on the increase of other oxidative enzyme and mitochondrial creatin kinase contents, prove this hypothesis [ ].
Mechanosensitivity in Cells and Tissues - NCBI Bookshelf
The gravitational field is the most constantly acting factor across the whole evolutionary development of all living organisms on Earth. Therefore, it is logical to suggest that mechanosensory mechanisms explained acts of the primary receptions of the mechanical force could be universal for the different cells. Indeed, they can be connected with the most ancient of cell structures, the membrane with the cortical cytoskeleton.
However, the means of forming a state of tension through the system of thin filaments is not yet clear enough. There are at least two possible ways of realising this function. The first being that actin forms a reticulum which is stiff enough to play the role of a skeleton. The second is through actin-myosin interactions, which are not exclusive to just the muscle cells.
The findings of Martens and Radmacher [ 10 ] on human fibroblasts showed that cell stiffness can be explained by the tension created by myosin transferring along the actin filaments. There is no doubt that external surroundings influence cell mechanical properties, most likely by means of reorganisation of the actions of the cytoskeleton. The data on cells cultured on different substrates proves this suggestion.
Both the topography of the substrate and its stiffness are of great importance. The data, obtained with the use of agents that damage the actin network meanwhile, show a leading role for the thin filament system in forming the cell structure and its rapid responses to changes in external conditions. Additionally, it was shown that different pathologic processes, especially malignant transformation of cells, can cause changes in the mechanical properties that tend to increase cell stiffness.
Evidently, this is also connected with the impaired regulation of the actin cytoskeleton state that takes place during the increase in cell proliferation speed. On the other hand, aging of the organism causes decreases in the stiffness of some cells that can be also connected with a change in cytoskeleton state.
There was a problem providing the content you requested
A change in the mechanical conditions also causes a change in cell stiffness, in particular in skeletal muscle fibres and cardiomyocytes. The decrease in the external mechanical force causes a decrease in the stiffness, and an increase, which in itself is correlated to the dynamics of nonmuscle actin isoform beta- and gamma- contents in the membrane fraction of muscle cells, is connected with differently directed changes in alpha-actinin-1 and alpha-actinin-4 contents in the cytoplasmic and membrane protein fractions.
A decrease in the force does not require a developed cytoskeleton, and as a result the cell stiffness should decrease. Thus, by summarising the data, we can suggest Figures 2 and 3 that a decrease in mechanical load causes the decrease in skeletal muscles fibre stiffness, likely due to the decrease in nonmuscle actin isoforms and alpha-actinin content in the membrane protein fraction, and also the decrease in cell respiration intensity that takes place in the skeletal muscles. At the increase of mechanical load on cells, in particular on the cardiomyocytes, during antiorthostatic disuse in rats, causes an intensification of the processes of cellular respiration, an increase of nonmuscle actin and alpha-actinin content in the membrane protein fraction, and an increase in cell stiffness also.
Therefore, there are differently directed changes in the alpha-actinin-1 and alpha-actinin-4 content in the cytoplasmic protein fractions. The results discussed here suggest several approaches to the regulation of cell mechanosensitivity and present a range of possibilities for prevention of the negative effects of microgravity and correction of negative changes connected with hypogravity syndrome. For example, the use of gene constructs or any preparations which provide stabilisation of the cortical cytoskeleton could prevent the negative effects of exposure to a zero-gravity environment.
However, the problem of perception of mechanical stimuli by different cells still requires further study. Indexed in Science Citation Index Expanded. Subscribe to Table of Contents Alerts. Table of Contents Alerts. Abstract This paper addressed the possible mechanisms of primary reception of a mechanical stimulus by different cells. Introduction The appearance of life on Earth and the evolution of all living organisms occurred under the influence of external physical fields, gravity, and electromagnetic fields.
Mechanical Properties of Cells 2. Cells Capable of Generating a Mechanical Force-Cardiomyocytes and Skeletal Muscle Fibres With the advent of atomic force microscopy, experimental studies on the mechanical properties of different cells were intensified [ 1 ]. Nonmuscle Cells Costa et al.
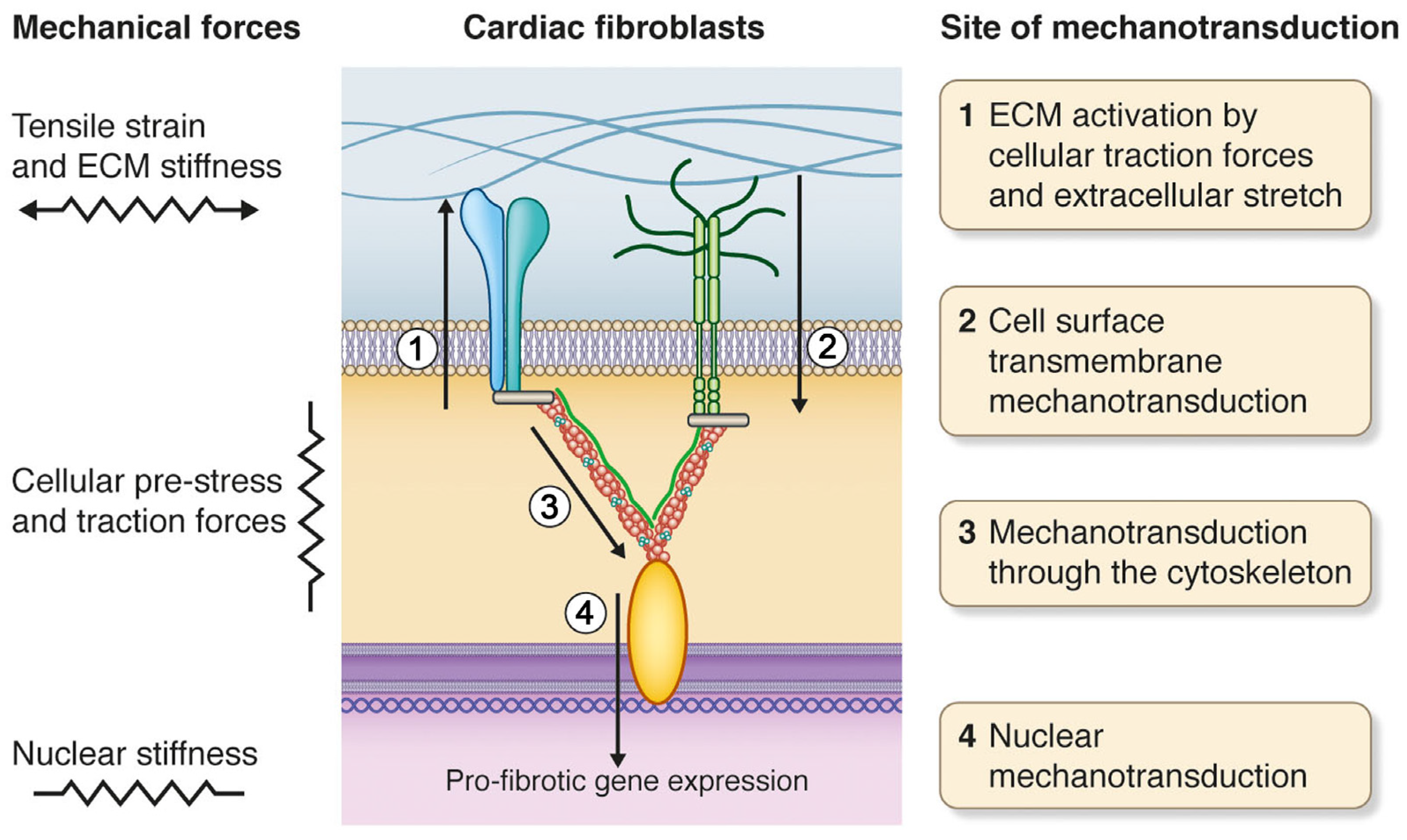
The Magnitude of Force Capable of Causing Cellular Responses The process of transformation of physical signals into biochemical ones, and the formation of appropriate cellular responses, is called mechanotransduction or mechanosensitivity [ 18 ]. Thus, a key question is what is the magnitude force that is able to cause a cellular response? Extracellular Matrix and Membrane Proteins It has been shown that the applying of stretching force to a culture of neurons or smooth muscle cells, through the extracellular matrix leads to an increase in the polymerisation of microtubules [ 30 , 31 ].
Mechanosensitive Ion Channels Mechanical stretching of cellular membranes, for example using the patch-clamp technique, changes the transportation activity of mechanosensitive ion channels as a result of conformational changes or changes in the lipid bilayer [ 33 , 34 ], or in the gate domains of the channel itself [ 35 , 36 ].
Cortical Cytoskeleton Today the role of cortical cytoskeleton in the regulation of ion channels is quite well established. Intracellular Structures It is well known that the action of external forces can lead to changes in the levels of gene expression. Changes in the external mechanical load cause a change in the internal mechanical tension of the cell and its deformation. Deformations can occur in the ion channels causing changes in their permeability for different ions, for example for calcium, which is a secondary messenger and can activate some signalling pathways.
Moreover, deformations can also occur in the cytoskeleton, both in the sarcomere for muscle cells and cortical cytoskeleton, causing the release of different signalling molecules and activation of downstream signalling pathways. Hypothetical mechanism for earlier cellular responses to changes in mechanical conditions. The principal difference between stretch and compression is characterized by the dissociation of different molecules from the cortical cytoskeleton, for example, alpha-actinin-1 at stretching and alpha-actinin-4 at compression.
Under cell stretch, there are cortical cytoskeleton deformations and subsequent shifts in actin filaments relative to each other in the stress fibres. This increases the probability of dissociation of the proteins that connect actin filaments, for example, alpha-actinin Under cell compression, this happens predominantly via membrane deformation so that the conformation of the alpha-actinin-4 binding sites e. This will lead to a release of proteins which connect with the membrane, for example, alpha-actinin The release of alpha-actinin-4 causes the activation of the expression of the alpha-actinin-1 gene and repression of own expression.
This occurs similarly for alpha-actinin The release of different proteins causes activation of different pathways and formation of the response to the increase or decrease in mechanical load. The proposed mechanism is only hypothetical and therefore needs to be checked experimentally.
Hypothetical mechanism for the development of adaptive responses of skeletal muscle fibres and cardiomyocytes to the change in mechanical conditions. Hypothetical links are shown by dashed arrowheads and contours. View at Google Scholar I. Herbert Wu, and C. View at Google Scholar T. View at Google Scholar E. View at Scopus D. Mechanical stress can modulate physiological processes at the molecular, cellular, and systemic level.
The primary target for mechanical stimulation is the plasma membrane of the cell, which can respond to variable physical stress with changes of the open probability of mechanosensitive ion channels. Thus, acting on ion channels in the plasma membrane, mechanical stress can elicit a multitude of biochemical processes — both transient and long-lasting — inside a cell. This may ultimately influence the function of tissues and organs in health and disease. Several stretch-induced signaling cascades have been described with multiple levels of crosstalk between the different pathways.
Increased sensitivity of the cells to mechanical stress is found under various pathological conditions. A detailed study of the underlying mechanisms may therefore help to identify novel therapeutic targets for a future clinical use. Turn recording back on. National Center for Biotechnology Information , U. Mechanosensitivity in Cells and Tissues Editors: For more information, see the Bookshelf Copyright Notice.
Rosen, and Leonid V. Mario Pellegrino, Barbara Calabrese, M.